Дискретная математика:
логика, группы, графы, фракталыАкимов О.Е.
4.7. Динамика популяций
Сейчас мы подойдем к вопросам, рассмотренным в предыдущем параграфе, с другой стороны. С этой целью обратимся к истории. Томас Мальтус (1766—1834) в 1798 г. опубликовал труд «Опыт о законе народонаселения», где утверждал, что население земли размножается в геометрической прогрессии, а средства его существования растут в арифметической прогрессии. Следовательно, говорил он, перенаселение земли будет неизбежным, что вызовет, в свою очередь, распространение нищеты, голода, болезней и ожесточенных войн. Выживут только те, кто пройдет этот «естественный отбор». Учение Чарлза Дарвина (1809 — 1882) о борьбе за существование есть прямая экстраполяция теории Мальтуса на весь органический мир. В начале октября 1836 г. Дарвин вернулся из кругосветного путешествия на корабле «Бигль», откуда привез богатейшую коллекцию растений и животных, а также бесценный «Дневник изысканий». Ровно через два года он прочел книгу Мальтуса о народонаселении. В течение 20 лет Дарвин упорядочивал и согласовывал добытый им материал с идеями Мальтуса. Наконец, 24 ноября 1859 г. вышел его знаменитый труд «Происхождение видов путем естественного отбора», где в третьей главе «Борьба за существование», в параграфе «Геометрическая прогрессия возрастания численности» он писал:
«Борьба за существование неизбежно вытекает из большой скорости, с которой все органические существа имеют тенденцию увеличивать свою численность. Каждое существо, в течение своей жизни производящее несколько яиц или семян, должно подвергаться уничтожению в каком-нибудь возрасте своей жизни, в какое-нибудь время года или, наконец, в определенные годы, иначе, в силу принципа возрастания в геометрической прогрессии, численность его быстро достигла бы таких огромных размеров, что ни одна страна не могла бы вместить его потомство. Поэтому, так как производится больше особей, чем может выжить, в каждом случае должна вестись борьба за существование либо между особями того же вида, либо между особями различных видов, либо с физическими условиями жизни. Это — учение Мальтуса, с еще большей силой примененное ко всему животному и растительному миру, так как здесь невозможно ни искусственное увеличение пищи, ни благоразумное воздержание от брака. Хотя в настоящее время численность некоторых видов и может увеличиваться более или менее быстро, но для всех видов это невозможно, так как земля не вместила бы их».Столкновение интересов и конкуренция за материальные, финансовые, людские ресурсы и рынки сбыта — это борьба за существование в сфере экономики. Следовательно, динамические законы, выведенные на примере роста какого-нибудь пенициллинового грибка затем можно будет распространить на широкий круг биологических, социальных и экономический явлений. Пусть x(t) — число особей грибковой популяции на момент времени t. Прирост особей за период Δt равен Δx = αxΔt, где α — коэффициент, характеризующий превышение рождаемости над смертностью (его еще называют удельной скоростью роста биологической массы). Отсюда вытекает экспоненциальный закон роста численности грибковой популяции, так как
,
,
,
ln |x| = αt + C, C = ln |x0| = αt0, x(t) = x0 exp [α(t – t0)].
Этот закон распространяется на любой биологический вид x, где отсутствует конкуренция. В случае, когда сосуществуют два вида x и y, возникает насильственная убыль особей x за счет уничтожения их особями y на величину βxyΔt, где β — коэффициент межвидовой конкуренции. В самом деле, с одной стороны, чем больше особей x, тем чаще жертва (x) будет попадаться хищнику (y), с другой стороны, чем больше хищников y, тем интенсивнее истребление жертв x. Но если имеет место конкуренция среди особей одного вида, то убыль определяется квадратичной зависимостью βx2Δt. Поэтому исходное уравнение изменится на уравнение, которое впервые было получено немецким биологом Ферхюльстом в 1845 году: Δx = αxΔt – βx2Δt и
или
При t → ∞, x → α/β, т.е. численность любой популяции стремится к постоянному значению. Если t не намного отличается от t0, т.е. когда популяция еще молода и конкуренция отсутствует, то имеет место первая формула для x(t) (ее можно получить, умножив числитель и знаменатель на exp [α(t – t0)] ).
В случае, когда жертва x и хищник y являются двумя различными видами, то имеет место система двух уравнений:
или
Здесь предполагается, что популяция-жертва x(t) является единственной пищей для популяции-хищника y(t). Прирост Δy за время Δt складывается из прибыли, обусловленной рождаемостью γxyΔt , и убыли из-за естественной смертности δyΔt, при этом γ — коэффициент рождаемости хищника, δ — коэффициент его смертности. Модель из двух функций x(t) и y(t), удовлетворяющих системе уравнений жертва-хищник, впервые были исследованы в работе Вольтерра 1931 года. Он выяснил, что с какой бы численности жертвы и хищники ни стартовали — M(x0, y0) или M'(x'0, y'0) — эта динамическая система всякий раз будет возвращаться к этим же начальным условиям, пройдя определенный замкнутый цикл, который показан на рис. 4.34а.
а
б
Рис. 4.34
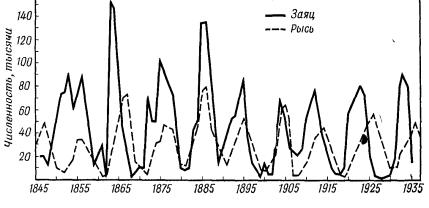
Классическим примером циклических изменений численности популяции являются колебания численности рысей и зайцев, приходящейся на определенную площадь соснового леса Канады. На рис. 4.34б показан график, на котором видно, что число особей популяции рыси достигает максимальной плотности примерно каждые 9,6 лет. Сходные колебания испытывает популяция зайца, причем максимальные значения его плотности наступали в среднем на год раньше , чем у рыси. Корреляция между численностью рыси и зайца подтверждает правильность модели Вольтерра жертва-хищник. Проследим циклическую динамику детально.
Если система жертва-хищник стартовала из точки M(x0, y0), то численность обоих видов начнет сокращаться до нижней точки 1 (рис. 4.34а), где хищников становится так мало, что зайцы начинают размножаться по законам, определяемым их собственными параметрами α и β. Увеличение численности зайцев приводит к увеличению численности рысей. Эта тенденция сохраняется до точки 2, после чего численность рысей продолжает увеличиваться, а численность зайцев начинает уменьшаться. В точке 3 наступает новая ситуация: хищников становится так много, что они поедают жертву еще до того, как она оставила потомство. Число зайцев стремительно убывает; возникает ситуация, когда рысь, не найдя добычи, погибает. На участке 3 – 4 наблюдается спад численности и хищника, и жертвы. На участке 4 – 1 спад численности рысей по инерции продолжается, но численность зайцев постепенно начинает восстанавливаться до величины, с которой начался цикл.
Многие авторы колебания численности популяций ставят в прямую зависимость от циклической активности Солнца. Здесь возникает множество спорных моментов. Уильямсон, приведя графики (рис. 4.35а) числа шкур рысей, заготовленные компанией Гудзонова залива в бассейне реки Маккензи в течение 100 лет (верхняя кривая), и числа солнечных пятен (нижняя кривая), заявил, что между этими циклами никакой связи нет, поскольку в начале XIX века пики совпадали по фазе, затем они шли в противофазе, потом снова синфазно. Действительно, на 11 пиков в численности рысей (точнее, числа шкур убитых рысей) приходится 10 пиков солнечной активности. Однако нельзя полностью исключить влияние Солнца на колебание численности популяций. Радиация заметно влияет на состояние живых организмов — это известный биологический факт, поэтому отсутствие частотного резонанса на всем рассматриваемом отрезке времени еще не означает отсутствие влияния солнечной активности на динамику популяций в более короткие периоды.
а
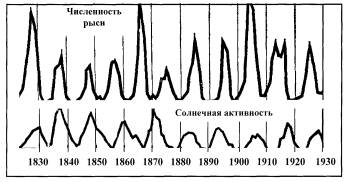
бРис. 4.35
Биоритмы бывают экзогенными, вызванными действием внешней среды (в первую очередь, суточной и сезонной периодичностью), и эндогенными, вызванными строением клеток, тканей, организма в целом, а также коллективным поведением особей популяции. Очень многие процессы в живой и неживой природе развиваются циклически, однако строгую математическую зависимость между этими колебаниями установить зачастую очень сложно, так как простые экзогенные колебания накладываясь на непростые эндогенные дают еще более сложную картину суммарных колебаний. Не удается даже точно предсказать пики морских приливов и отливов, хотя эта механическая задача несравнимо проще любой биологической. Дальнейший анализ этого вопроса показывает, что циклический закон Вольтерры не всегда доминирует в динамике популяций. Статистические данные по рысям и зайцам — это, пожалуй, единственный пример, где прослеживается долговременная периодичность численности особей. Чаще всего колебания носят кратковременный периодический характер, затем следует сбой, флуктуация колебаний или хаотические количественные всплески, поскольку на численности потомства сказываются и холодная зима, и засушливое лето, и сотни других факторов, среди которых солнечная активность будет не на последнем месте.
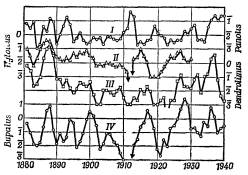
На рис. 4.35б показаны колебания численности популяций куколок Panolis (I), куколок Hyloicus (II), зимующих личинок Dendrolimus (III) и куколок Bupalus в Лецлингере (IV), в штуках на 1 м2 (в логарифмическом масштабе). Такой вид кривых плотности заселения имеет большинство популяций. Тем не менее модель Вольтерры является образцом конструктивного подхода в сфере сложных динамических систем, в которых наблюдаются автоколебания.
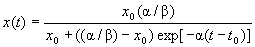
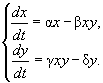
|
|